The Maximum Genetic Diversity (MGD) Theory - The Golden Gnomon
The Maximum Genetic Diversity Theory of Evolution
Overview
This is the official website for Professor Shi Huang and the Maximum Genetic Diversity (MGD) Theory. This site explores the groundbreaking discovery of the Genetic Equidistance Phenomenon (GEP) and its implications for human origins, including the revolutionary Out of East Asia Theory as an alternative to the traditional Out of Africa model. Our mission is to explain, document, and share the evidence for MGD — a framework for understanding biodiversity, human origins, and molecular evolution.
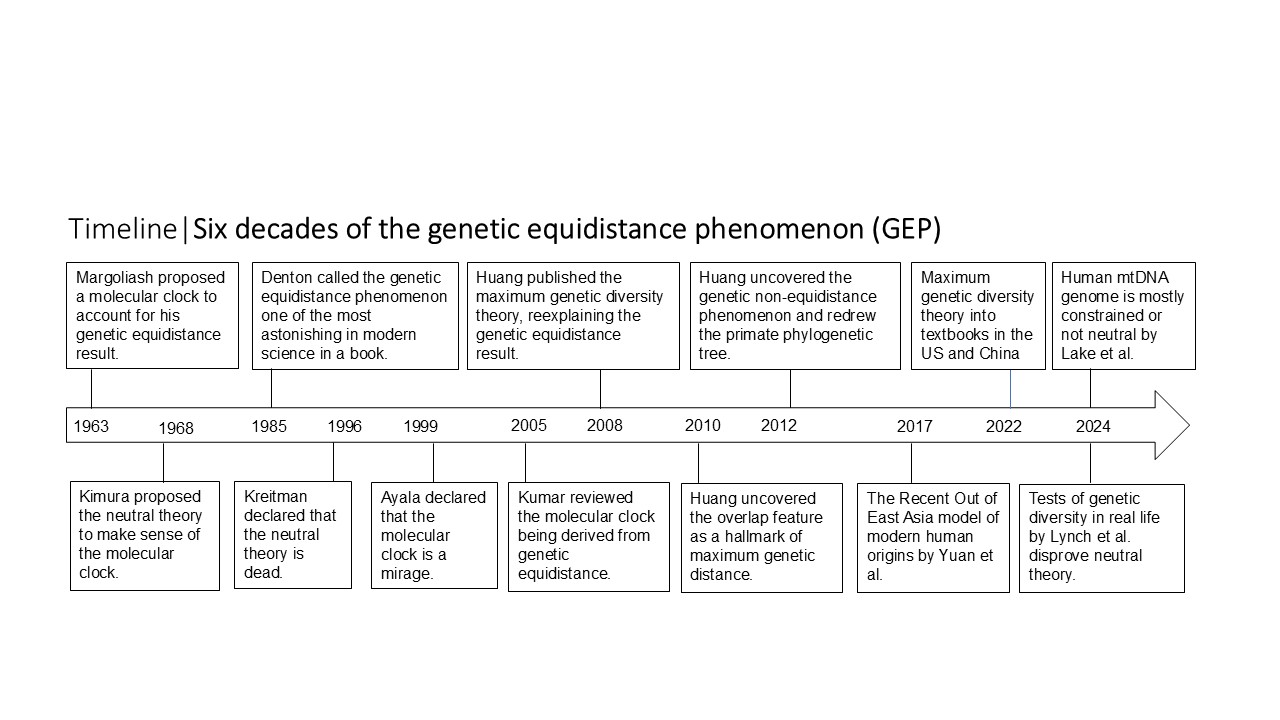
The MGD theory was inspired by an independent rediscovery of the GEP, first described in 1963. GEP originally led to the strict molecular clock (SMC)hypothesis, which in turn gave rise to the neutral theory. However, the SMC has since been disproven and replaced by the “relaxed clock,” which allows different species to have different mutation rates. This revision undermines the neutral theory—the only premise that supported the strict clock model. With the neutral theory effectively invalid, the MGD theory stands as the sole remaining explanation for genetic equidistance. A distance independent of rate can only reflect a saturation limit: the maximum genetic distance.
Explaining GEP is therefore a strict binary choice: either the SMC and neutral theory hold—excluding MGD—or they fail, leaving MGD as the only viable explanation. This establishes a clear “either–or” relationship. The competition between MGD and the Neutral Theory is an absolutely unambiguous binary choice—not only by logic but also by fact. For 60 years, molecular evolution has had only two competing explanations for the genetic equidistance phenomenon, and no others. This stands in sharp contrast to Darwin’s theory of natural selection, where countless rival theories exist—without exaggeration.
The MGD theory overturns many phylogenetic conclusions that relied on the now-discredited SMC and neutral theory. It redefines our understanding of evolutionary change by emphasizing the saturation limits of genetic diversity. Importantly, it also challenges prevailing models of human origins, providing strong support for the Out of East Asia hypothesis for the origin of modern humans or Homo sapiens.
In geometry, a gnomon is a figure that, when combined with an existing shape, produces a new figure similar to the original. This concept was introduced in Euclid’s Elements (Book II, Definition 2) and has been generalized to describe any rule of transformation that generates patterns that are both distinct and self-similar. The golden gnomon refers to the gnomon of the golden triangle — a metaphor for the iterative, self-similar process of creation in evolution. Our goal in evolutionary biology is to uncover the “gnomon” that drives the endless generation of life’s diversity and complexity/order.
MGD Concepts
Explore the foundational concepts of the Maximum Genetic Diversity (MGD) theory:
Concepts
GEP (Genetic Equidistance Phenomenon):
A simpler species is equally distantly related to two or more complex or equally complex species in a sequence alignment when the genetic distance is measured by the identity matrix or percentage non-identity.
MGD (Maximum Genetic Diversity):
The upper limit in genetic distance or diversity, as measured by the identity matrix or percentage differences, that can be reached during evolution over long periods. The new concept of maximum genetic diversity is mainly in regard to the form of genetic diversity as measured by genetic distance.
Microevolution:
In the MGD theory, the term encompasses evolutionary changes both within a species and between species over both short and long periods, as long as these changes do not involve major epigenetic modifications.
Macroevolution:
In the MGD theory, the term describes new species formation that involves both genetic and epigenetic changes, particularly involving increases in epigenetic complexity.
Axiom 1
The more complex the phenotype, the greater the restriction on the choice of molecular building blocks. Complex/ordered systems need higher precision building parts. In biology, this means there is an inverse relationship between genetic diversity and epigenetic complexity. Complexity is defined as the number of cell types.
Axiom 2
For any system that can allow a limited level of random errors or noises in molecular building parts, such errors may be beneficial, deleterious, or neutral depending on circumstances.
Limited errors at optimum level are more likely to be beneficial than deleterious because they are, after all, within tolerable levels and confer economy in construction and the strongest possible adaptive capacity or robustness to environmental challenges.
In biology, substituting “errors in building blocks” with “genetic diversity” gives the equivalent concept. Axiom 2 in fact highlights the valid parts of Kimura’s and Darwin’s theories.
Axiom 3
Matter or randomness and consciousness or cognition are opposite to each other. High randomness inside the body of an individual must result in poor mental function, and the measure of randomness is genetic diversity, as diversity originates from random mutational events. Thus, complex species with higher cognitive capacity must have lower randomness or genetic diversity.
Axiom 4
Deleterious mutations, which impair a trait, can be mitigated by compensatory mutations. Populations with high genetic diversity are better equipped to tolerate or counteract harmful mutations, making it challenging for natural selection to maintain trait quality and eliminate detrimental variants. Therefore, reducing genetic diversity may be necessary to preserve high-quality traits and effectively remove harmful mutations.
Core Principles of the MGD Theory
Maximum genetic diversity tends to be higher for simpler taxa and lower for more complex taxa.
Macroevolution involves changes in organismal complexity and often results in an increase in complexity, which is mirrored by an increase in the precision of the building parts or a decrease in the allowed range of the standard deviations for the parts.
Microevolution, on the other hand, is an increase in genetic diversity within the allowed standard deviation ranges without significant change in complexity. It encompasses both evolution within species and evolution from one species to another.
At the saturation phase, microevolution involves turnover of alleles at the equilibrium level of genetic diversity.
The positions that are conserved in simpler taxa tend to also be conserved in more complex taxa. The positions that are free to change in more complex taxa tend to also be free to change in simpler taxa.
Genetic distance among taxa and genetic diversity within a taxon is mostly at the optimum level today after a very long evolutionary time, especially so for fast-evolving sequences. Any level higher or lower than the optimum would be negatively selected.
As genetic diversity, so long as it is within the maximum level, facilitates adaptation, it would be positively selected to quickly reach the optimum level. The optimum concept here means a Pareto optimum or simply the best that can be achieved due to a balance between positive and negative selection at a particular time point under a specific level of epigenetic or organismal complexity.
As time, environments, and complexity change, the optimum level of nucleotide diversity will also change.
Genetic variants are mostly functional or under balancing selection and quasi-neutral (under both positive and negative selection) rather than neutral.
Genetic distance or molecular distance between two taxa of different complexity is not contributed equally by mutations in the two lineages but rather is mostly contributed by mutations in the simpler lineage.
Non-conservation is not non-function. Fast-changing non-conserved sequences play more important roles in adaptation to the environment than the slowly changing conserved housekeeping genes.
Lower MGD means higher homozygosity, which is however very different from the higher homozygosity due to inbreeding. Lower MGD results in higher fitness traits because there are more common alleles or good alleles becoming homozygous.
In contrast, inbreeding leads to lower fitness traits (inbreeding depression) due to homozygosity in minor alleles or deleterious alleles. Inbreeding shows long runs of homozygosity (ROH) but lower MGD does not.
The origin of the first life involves a reduction in the randomness of the life-building molecules, which is fundamentally similar to the reduction in genetic diversity or randomness during the step-wise increase in complexity in macroevolution. They all involve the same complexifying force or anti-randomness force.
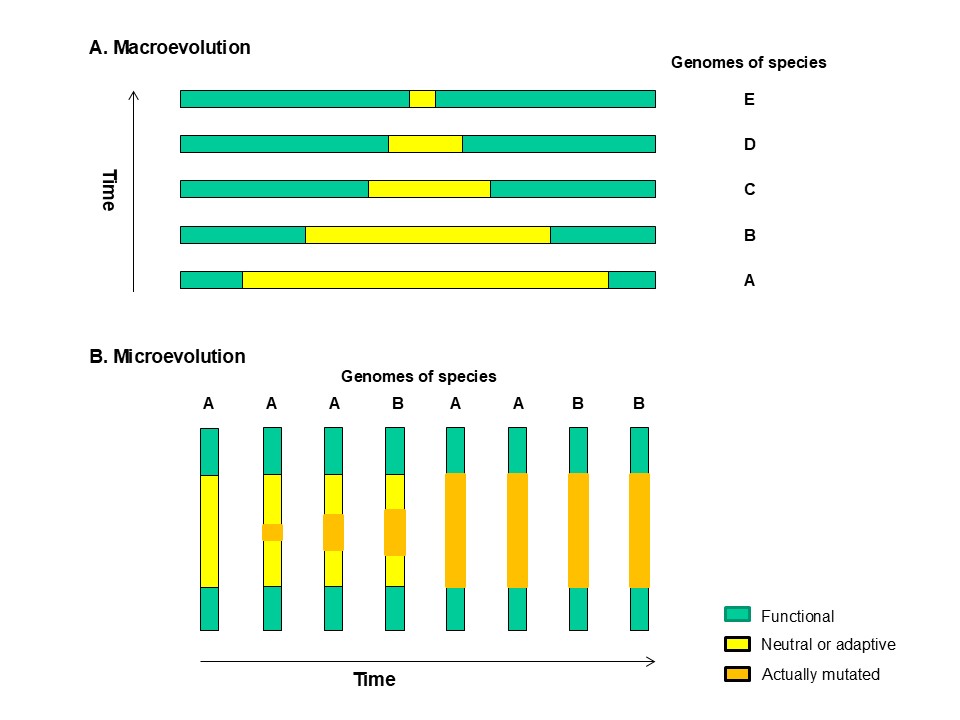
Schematic representation of the maximum genetic diversity theory of evolution.
A. Macroevolution.
As species evolve from simple to complex (taxon A → taxon E), the maximum level of genetic diversity that a taxon can tolerate is reduced.
B. Microevolution.
Accumulation of random mutations within the tolerated range of genetic diversity leads to speciation (from taxon A → taxon B) without large changes in epigenetic complexity or in the maximum genetic diversity that the taxon can tolerate.
GEP reinterpreted by MGD
The MGD theory explains the GEP as a result of maximum genetic distance. Over a long evolutionary time and for fast-evolving DNAs, the genetic distance between species has reached the maximum level.
The distance between the ingroup species and a simpler outgroup taxon is mainly determined by the maximum genetic diversity of the simpler outgroup. This distance is equal to the maximum distance allowed within members of the simpler outgroup, e.g., the distance between humans and fishes equals the maximum distance between different taxa of fishes.
Changes in the lineage leading to the simpler outgroup mask any changes in the lineages leading to the ingroup taxa.
There are in fact two kinds of genetic equidistance results. For long evolutionary timescale or for fast-evolving sequences, one would observe “maximum genetic equidistance”: different species are equidistant to a species of lower or equal complexity. The original result of Margoliash is maximum genetic equidistance.
For short evolutionary timescale or for slow evolving sequences, one observes “linear genetic equidistance” where the molecular clock holds and the distance is still linearly related to time: when ingroup species have similar mutation rates, they would be equidistant to a lower or equal complexity outgroup.
This explanation of the genetic equidistance result by the MGD theory can also be easily illustrated by a simple thought experiment. If we can create a yeast, a fish, and a human being by using identical genes for their shared homologs and let the three organisms diverge for an infinite amount of time or about 500 million years with each organism remains phenotypical largely the same as today, a gene in yeast would have changed a lot to a maximum of, say, 50%, while its homolog in fish would have changed to a maximum of, say, 30%, and its homolog in human would have changed very little, say less than 1%.
Any more changes than 50% would be lethal to yeast; any more changes than 30% would be lethal to fishes; and any more changes than 1% would be lethal to humans. The reason that a gene in yeast can change much more than in fish, which is still more than in human, is because a gene in human encounters far more functional constraints than its homolog in fish or in yeast.
Thus the genetic distance between yeast and human or fish is mainly determined by the mutations in yeast. In this case, the 50% change in yeast would account for the genetic distance of 50% identity between yeast and human or between yeast and fish, as well as 50% identity in within species distance in yeast.
The 30% change in fish would account for the genetic distance of 30% identity between fish and human. In contrast, the neutral theory would predict that both human and fish can also, like yeast, change up to 50% or more and would have a genetic distance of 50% identity.
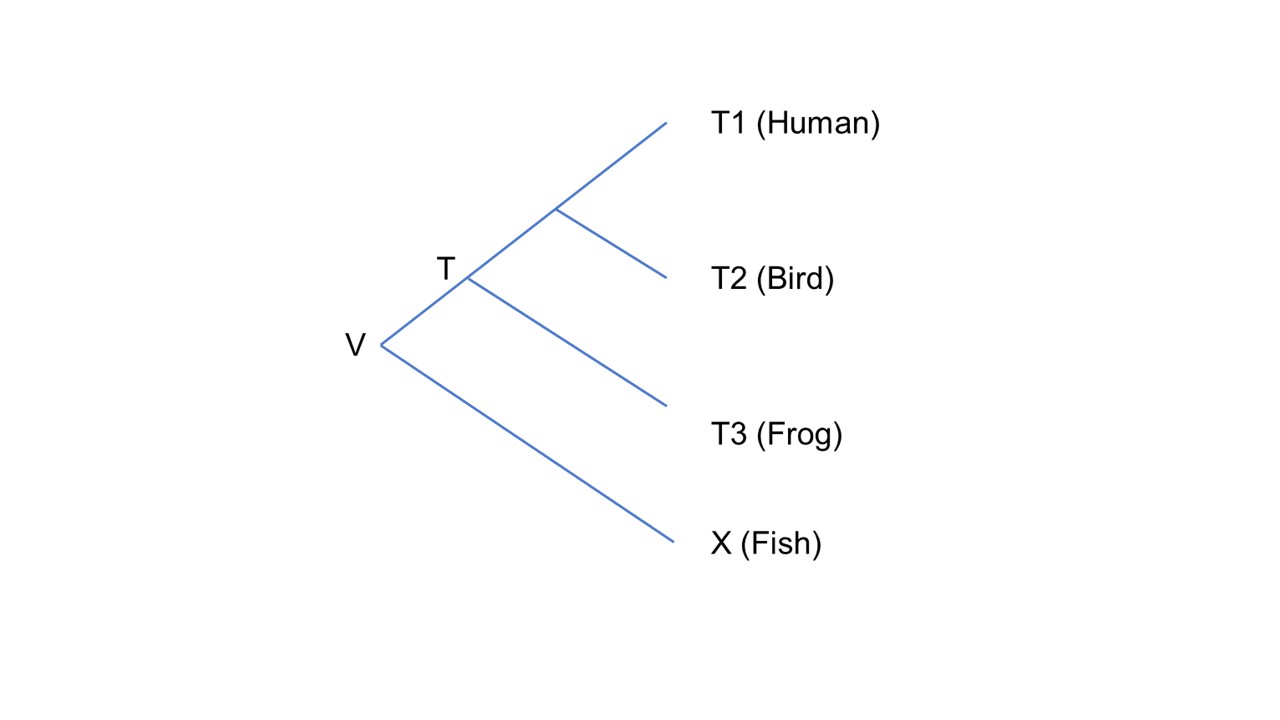
Figure. Illustration of the genetic equidistance phenomenon and its interpretation known as the molecular clock.
Comparison for tetrapod species (T1–T3; human, bird, frog), which are known to have a most recent common ancestor (T), and another species (X; fish). Time flows from the past (left) to the present (right).
Species X is the outgroup species and is equally distant to species T1–T3, the ingroup species, both in terms of time of separation and sequence difference, as measured by the identity matrix.
This is illustrated by the lines linking X to T1, T2, and T3 being of equal length, indicating the same level of divergence in both time and sequence.
The GEP refers to the fact that X is equally different in protein sequence to T1, T2, and T3. Evolutionary lineages leading to species T1–T3 separated from the lineage leading to X at the same point, V.
Furthermore, species T1–T3 are products of an evolutionary process that has been ongoing for the same duration since their common ancestor, V. Therefore, if a given protein exhibits equal divergence when comparing the same fish protein with proteins from different tetrapods, it suggests that the rate at which differences accumulate is similar among tetrapods (T1–T3).
The implicit assumption here is that the molecular distance among the species has not yet reached its maximum level, allowing us to infer the rate at which sequence differences accumulate. If, however, the distance has reached an upper limit, it would no longer be related to time or mutation rate, rendering the inference of the rate invalid.
Evidence
Scientific evidence supporting MGD:
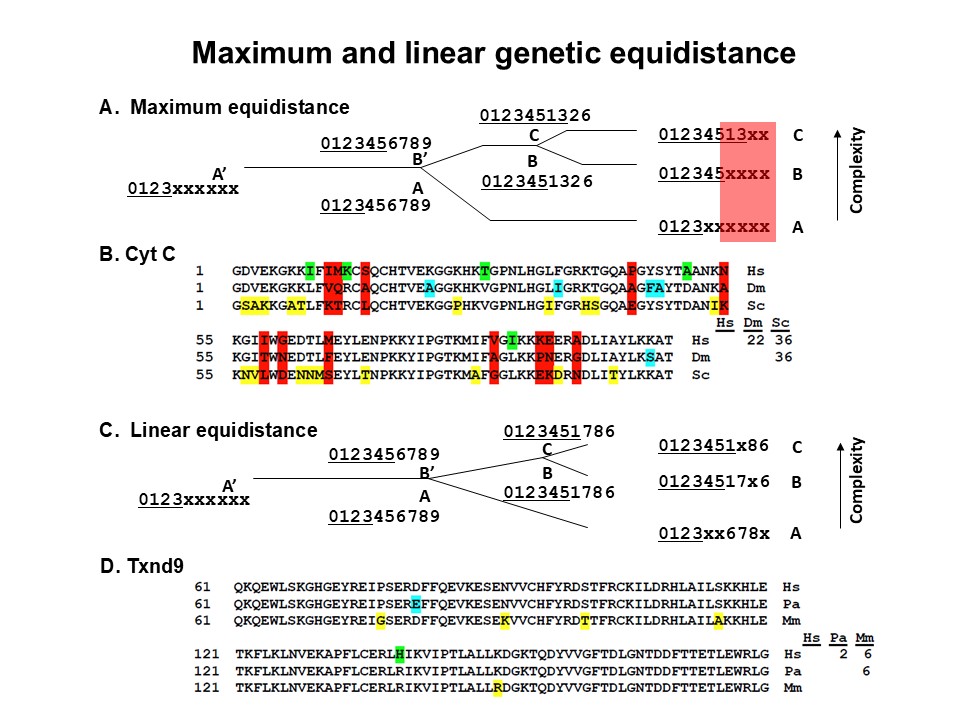
The overlap feature: There are three ways to test which of the two different interpretations of genetic equidistance is true, the molecular clock versus the MGD theory. When aligning a protein sequence from three taxa, the conserved positions would show identical amino acids for all three taxa. The non-conserved positions can be categorized into two types. One type involves mutation in only one of the three taxa, while the other two taxa retain either identical amino acids or remain unchanged. The second type consists of positions where each taxon has a unique amino acid at the same location (the red colored positions in the Figure below). This indicates that at least two taxa had gone through a unique mutation at that same position, and hence their mutations overlapped at that position. Such positions are referred to as overlap positions. Since one mutation is enough to lead to a difference of one between two taxa as measured by identity matrix, overlapped mutations at the same position do not increase molecular distances and are hence hallmarks of mutation saturation. At maximum saturation, each free-to-change position in a taxon would have mutated, resulting in a higher fraction of overlap positions among the non-conserved positions than expected by chance. This is termed the high overlap ratio (HOR) phenomenon.
Figure. The genetic equidistance result and the maximum genetic diversity theory. A. Maximum genetic equidistance. A ten amino acid peptide is used to illustrate the evolution process. When the protein is fast evolving, the observed equidistance today would be maximum distance with a high overlap ratio. The figure shows 4 overlap positions with an overlap ratio 1. The distance of C-A is 60%, the same as that of B-A. This is a schematic representation of the original Margoliash genetic equidistance result. B. An example of maximum genetic equidistance. Alignment of human, drosophila, and yeast cytochrome C proteins. Human differs from drosophila in 22 amino acid positions. Human and drosophila are equidistant to yeast with 36 amino acid differences. There are 12 overlap positions (in red color) and the overlap ratio is 12/22 = 55%. Other mutant positions are colored in green, blue and orange. C. Linear genetic equidistance. When the protein is slowly evolving, assuming molecular clock holds, the observed equidistance today would be linear distance with a low overlap ratio. Here every substitution in any species would mean an increase in distance. The figure shows 0 overlap position with an overlap ratio 0. The distance of C-A is 50% and equals that of B-A. D. An example of linear equidistance. Human, orangutan, and mouse TXND9 gene alignment. There are 2 amino acid differences between human and orangutan, which are equidistant to mouse with 6 amino acid differences. The overlap ratio is 0/2 = 0.
Genetic non-equidistance: The second way to test the molecular clock versus the MGD theory is the genetic non-equidistance result despite equidistance in time. The MGD theory predicts that maximum equidistance would only result when the outgroup is less complex than the sister species. If the outgroup is more complex, then its maximum distance with the ingroup sister species would be determined by the MGD of each of the ingroup species, which may not be the same for all the ingroup species. However, the molecular clock would predict genetic equidistance to the outgroup regardless if the outgroup taxon is more or less complex. For example, human is the outgroup to the Sauropsida clade containing snake and bird. The molecular clock predicts that humans should be equidistant to snakes and birds in protein sequence. However, the MGD theory predicts that birds should be closer to humans than snakes should because birds should be more complex than snakes.
The actual data validated the MGD theory. Genetic non-equidistance to humans despite equidistance in time has also been found for sister species within the teleost fish clade, the arthropod phylum, the Porifera phylum, and the fungi kingdom. In all five cases where the difference in complexity of the ingroup sister species can be intuitively inferred (octopus vs. cockle, terebratulina vs. lingula, bird vs. snake, dragonfly vs. louse, and smut vs. yeast), the more complex species always shows greater sequence similarity to humans in fast-evolving genes, fully conforming to the predictions of the MGD theory but not that of the molecular clock. It has recently been shown that octopus indeed has the lowest heterozygosity level among mollusks, and that the highest heterozygosity level among mollusks is found for the least complex solenogastres. Also, by whole genome sequencing analysis, two new world monkeys are found to be non-equidistant in nucleotide sequence to humans with the most primitive monkey marmoset to be more distant to humans than the owl monkey.
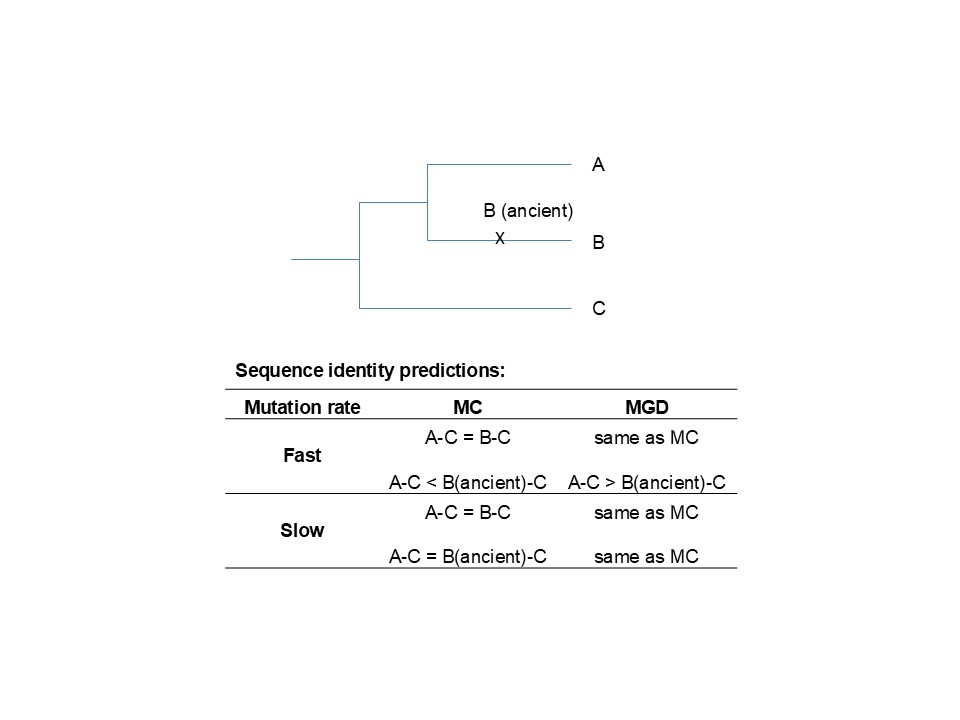
Ancient DNA/proteins: The third way to test the molecular clock versus the MGD theory is to make use of ancient DNA/proteins. Under the molecular clock (MC) and neutral theory, an ancient species should exhibit less sequence divergence from an outgroup than its extant sister species. However, the MGD theory posits that ancient DNA should be more divergent from the outgroup than its extant sister species.
Consider three extant species of similar phenotypic complexity: A, B, and C, where A and B are sister ingroup species, and C is the outgroup. According to the GEP and MGD theory, A and B are equidistant to C, with the distance at an upper limit for fast-evolving genes. Due to differing environmental conditions, ancient species would share variants adapted to past environments, which would have been replaced in their extant lineages by alleles suited to current environments. Consequently, the genetic identity between B and C (or A and C, as BC = AC per the GEP) would be greater than that between ancient B and extant C. However, for slowly-evolving genes and for an ancient taxon from the relatively recent past, the B-C identity is expected to be comparable to the B(ancient)-C identity, as significant evolutionary changes are unlikely to accumulate over a relatively short period.
Figure. Schemes for testing the molecular clock (MC) hypothesis vs. the MGD theory by using ancient DNA. Extant species are represented by A, B, and C. Ancient taxon from the relatively recent past is represented by B (ancient).
Dead neutral theory: If the strict molecular clock and neutral theory are wrong, then the Maximum Genetic Diversity (MGD) theory must be correct or provides the correct explanation for the genetic equidistance phenomenon. Many may think that falsifying the neutral theory does not automatically validate MGD—but in fact, the logic is decisive.
For two sister species derived from a common ancestor, their genetic distance increases over time according to mutation rate and time. If they are equidistant in sequence identity to an outgroup species, and mutation rates are equal, this equidistance could still reflect time-dependent divergence (but not necessarily). However, if mutation rates differ—that is, if the strict molecular clock and neutral theory fail—then the observed equidistance cannot be explained by mutation rates. A distance independent of rate can only reflect a saturation limit: the maximum genetic distance.
Thus, explaining genetic equidistance is a strict binary choice: either the strict molecular clock and neutral theory hold, excluding MGD, or they fail, leaving MGD as the only viable explanation. This creates a clear “either-or” relationship. The competition between MGD and the Neutral Theory is an absolutely unambiguous binary choice—not only by logic but also by fact. For 60 years, molecular evolution has had only two competing explanations for the genetic equidistance phenomenon, and no others. This stands in sharp contrast to Darwin’s theory of natural selection, where countless rival theories exist—without exaggeration.
This reasoning is both simple and intuitive, making the inevitability of MGD evident even to non-specialists. The strict molecular clock has long been abandoned, replaced by the “relaxed molecular clock,” which undermines the neutral theory, the only premise supporting strict clock predictions. With neutral theory effectively invalid, the MGD theory emerges as the sole remaining explanation for genetic equidistance.
Applications
The maximum genetic diversity theory has been instrumental in directing productive research on both evolutionary problems and important biomedical problems. The theory does not mean discarding the old assumptions but merely making them more limited in their scopes. One must carefully select those DNAs that may follow those assumptions.
Phylogenetics
The maximum genetic diversity theory should help resolve difficult historical problems such as the phylogenetic tree of life. Past methods have no concept of maximum distance and use mostly non-informative distance data for inferring phylogeny. The slow clock method based on the MGD theory makes use of only slow-evolving sequences and thus ensures the linear relationship between distance and time. Its results therefore will be more objective and independent of the variations in sequence selections and investigators. The slow clock method has re-established a primate phylogeny that humans and pongids are two separate groups, which has long been the consensus view of paleoanthropologists.
To truly neutral sequences still at the linear phase of divergence, many of the assumptions of the neutral theory such as the infinite sites model would be valid. Thus phylogenetics research can largely proceed as before except that one now has a standard to separate the neutral from the noninformative DNAs. One must now distinguish two different kinds of high sequence similarity, one due to less time of separation and the other because of common construction resulting in using similar parts (convergent evolution).
Origins of Modern Humans — Recent Out of East Asia Theory
The out of Africa model of modern human origins is based on the molecular clock and the neutral theory. The high genetic diversity of Africans is interpreted to mean a deeper evolutionary time for Africans if one assumes the molecular clock. Also, the infinite site model is assumed in order to infer the derived allele status, which is critical for rooting the phylogenetic tree in Africans by using the outgroup rooting method. However, both of these assumptions are invalid according to the MGD theory and experimental data. By using informative variants and allowing recurrent and back mutations, we have built a new model of modern human origins, the Recent out of East Asia (ROE) model. The ROE model is consistent with the multiregional model in terms of autosomal evidence, which indicates that the major races have separated for ~2 million years as originally claimed by the multiregional model. However, uniparental DNA data indicates a single origin in East Asia at a more recent time.
The likely scenario is that modern humans first evolved in East Asia as marked by a new modern version of uniparental DNAs and then migrated to Europe and Africa and admixed with local less modern people. Admixture led to replacement of uniparental DNAs and autosomal DNAs so that Europeans or Africans would have modern uniparental DNAs but largely local autosomal DNAs. Ancient human DNA should be very informative in falsifying the incorrect models. Our analysis of ancient DNA samples has confirmed the ROE model. In contrast, researchers who believe in the out of Africa model have yet to report any ancient DNA evidence for their model but have instead found support for the ROE model, i.e., ancient DNA samples of 40,000–45,000 years old found in Europe and East Asia are East Asian-like rather than African-like.
Biomedical Problems — Complex Diseases and Traits
Most complex traits and diseases are partly inheritable and presumably caused by polymorphic genetic variations such as SNPs. The neutral theory views most such variations to be nonfunctional and neutral and hence the study of complex traits and diseases has in the past focused on searching for a few functional variants. Although such GWAS studies have met some successes in identifying a number of variants, these variants account for only a small fraction of the total trait variation and their functional roles typically remain unclear.
The maximum genetic diversity theory predicts that complex diseases may be caused by excess genetic noise over a threshold and may serve to prevent an infinite increase in genetic diversity. Complex traits evolved as a result of suppressing genetic noises and hence should be susceptible to damage by excess noises. Also, insufficient amount of genetic diversity may hurt adaptive capacities such as immunity. The quantitative variations in a complex trait may correlate with the number of genetic variations.
Results from our efforts in testing the MGD theory have shown the expected pattern that higher minor allele contents (MAC) or noises correlate with many complex diseases. These include association of MAC with higher lung cancer incidence in mice and humans. Also, Parkinson’s disease patients have higher MAC than controls and a selected set of ~37,000 minor alleles can predict 2% of Parkinson’s patients. Other diseases that show higher MAC include schizophrenia, type 1 diabetes, type 2 diabetes, lung cancer, and Alzheimer’s.
To directly examine the self-evident antagonistic relationship between cognition or consciousness and randomness or genetic diversity, we have performed a study analyzing the genotype and phenotype data from more than 400,000 people in the UK. We calculated multiple measures of genetic diversity for each individual, and examined which traits these measures were associated with using linear regression analysis that controlled for confounding factors. Among the 17 traits examined, only educational attainment, which is highly correlated with cognition or IQ, has the most robust relationship with genetic diversity, and it is an inverse association. This association is likely to be causal, since only the brain-expressed genes, but not the brain-non-expressed genes, showed an association. This result is likely to be free from the interference of confounding factors, because the correlation of non-synonymous variants is significantly higher than that of synonymous variants or intronic variants. Consistently, animal studies have also revealed an inverse relationship between learning and memory and genetic diversity.
Why Sex?
Almost all eukaryotes reproduce sexually, through meiosis which generates haploid gametes from a diploid cell. The purpose of sex has long remained a mystery. The common explanation is that sexual reproduction increases genetic diversity. However, asexual organisms such as bacteria generally have much higher genetic diversity than eukaryotes. There is also the suggestion that sexual reproduction can remove chromosomal and epigenetic abnormalities or other deleterious mutations. However, such abnormalities could also be removed by natural selection of abnormal phenotypes.
The MGD theory offers a straightforward solution to the mystery of sex. According to the theory, macroevolution from a simple taxon to a higher complexity taxon requires a reduction in genetic diversity (at the nucleotide level). The reduction in genetic diversity in an individual of the simple taxon is necessary for the individual to become the incipient individual of the more complex new taxon. As the overall level of genetic variation in an offspring is mostly determined by the inheritance of the combination of single nucleotide variants carried by the parents, sexual reproduction can either increase or decrease the genetic variation in an offspring relative to the parents, but asexual reproduction can only increase the genetic variation in an offspring. Thus, sexual reproduction is essential for reducing genetic diversity necessary for the macroevolution of higher complexity.
Publications
Explore all publications by Shi Huang on Google Scholar. Below is a comprehensive list of papers by Huang related to the Maximum Genetic Diversity (MGD) theory:
Huang, S. (2008) Histone methylation and the initiation of cancer. Cancer Epigenetics, Ed. Tollefsbol, T., CRC Books.
Huang, S. (2008) Ancient fossil specimens are genetically more distant to an outgroup than extant sister species are. Rivista di Biologia / Biology Forum, 101: 93-108.
Huang, S. (2008) The genetic equidistance result of molecular evolution is independent of mutation rates. J. Comp. Sci. Syst. Biol., 1: 92-102.
Huang, S. (2008) Modeling the creative process of the mind by prime numbers and a simple proof of the Riemann hypothesis. arXiv 0810.0095.
Huang, S. (2009) Molecular evidence for the hadrosaur B. canadensis as an outgroup to a clade containing the dinosaur T. rex and birds. Rivista di Biologia / Biology Forum, 102: 20-22.
Huang, S. (2010) The overlap feature of the genetic equidistance result, a fundamental biological phenomenon overlooked for nearly half of a century. Biological Theory, 5: 40-52.
Huang, S. (2010) Primate phylogeny: molecular evidence for a pongid clade excluding humans and a prosimian clade containing tarsiers. Nature Precedings; Link.
Yuan, D., Zhu, Z., and Huang, S. (2012) A more complete reinterpretation of molecular evolution nearly half of a century later supports the multiregional hypothesis rather than the out of Africa hypothesis. News magazine from the Chinese Academy of Social Sciences, Issue 389, 2012.12.07 (in Chinese).
Huang, S. (2012) Primate phylogeny: molecular evidence for a pongid clade excluding humans and a prosimian clade containing tarsiers. Sci China Life Sci, 55: 709-725.
Yuan, D., Zhu, Z., Tan, X., et al. (2012) Minor alleles of common SNPs quantitatively affect traits/diseases and are under both positive and negative selection. arXiv:1209.2911.
Hu, T., Long, M., Yuan, D., et al. (2013) The genetic equidistance result: misreading by the molecular clock and neutral theory and reinterpretation nearly half of a century later. Sci China Life Sci, 56: 254-261.
Huang, Y., Xia, M., and Huang, S. (2013) Evolutionary process as unveiled by the maximum genetic diversity hypothesis. Hereditas (Beijing), 35: 599-606.
Zhu, Z., Lu, Q., et al. (2013) Role of genetic polymorphisms in transgenerational inheritance of inherent as well as acquired traits in budding yeast. arXiv:1302.7276.
Yuan, D., Zhu, Z., Tan, X., et al. (2014) Scoring the collective effects of SNPs: association of minor alleles with complex traits in model organisms. Sci China Life Sci, 57: 876-888.
Huang, S. (2014) Human origins as told by informative DNAs. The development of Chinese Anthropology in the 21st century, Ed. H. Xi, W. Liu, Z. Chen, Intellectual Property Press, Beijing.
Zhu, Z. and Huang, S. (2014) Role of SNPs in complex diseases. J. Central South Univ., 39: 969-974.
Zhu, Z., Man, X., et al. (2015) Collective effects of SNPs on transgenerational inheritance in Caenorhabditis elegans and budding yeast. Genomics, 106: 23-29.
Zhu, Z., Lu, Q., et al. (2015) Enrichment of minor alleles of common SNPs and improved risk prediction for Parkinson's disease. PLoS One, DOI: 10.1371/journal.pone.0133421.
Zhu, Z., Lu, Q., et al. (2015) Collective effects of common SNPs in foraging decisions in Caenorhabditis elegans and an integrative method of identification of candidate genes. Sci. Rep., 5, 16904; doi: 10.1038/srep16904.
Zhu, Z., Lu, Q., et al. (2015) Compatibility between mitochondrial and nuclear genomes correlates with the quantitative trait of lifespan in Caenorhabditis elegans. Sci. Rep., 5, 17303; doi: 10.1038/srep17303.
Luo, D. and Huang, S. (2016) The genetic equidistance phenomenon at the proteomic level. Genomics, 108: 25-30. doi:10.1016/j.ygeno.2016.03.002.
Huang, S. (2016) New thoughts on an old riddle: what determines genetic diversity within and between species. Genomics, 108: 3-10. doi:10.1016/j.ygeno.2016.01.008.
Huang, S. (2016) Editorial: Special issue on the comprehensive functionality of genomic DNAs. Genomics, 108: 1-2. doi:10.1016/j.ygeno.2016.06.001.
Zhu, Z., Lu, X., et al. (2016) Close genetic relationships between a spousal pair with autism-affected children and high minor allele content in cases in autism-associated SNPs. Genomics, 109: 9-15.
Yuan, D., Lei, X., et al. (2017) Modern human origins: multiregional evolution of autosomes and East Asia origin of Y and mtDNA. bioRxiv. doi:10.1101/101410.
Yuan, D. and Huang, S. (2017) Genetic equidistance at the nucleotide level. Genomics, 109: 192-195.
He, P., Lei, X., et al. (2017) Accumulation of minor alleles and risk prediction in schizophrenia. Sci. Rep. doi:10.1038/s41598-017-12104-0.
Lei, X. and Huang, S. (2017) Enrichment of minor allele of SNPs and genetic prediction of type 2 diabetes risk in British population. PLoS ONE, 12(11): e0187644. https://doi.org/10.1371/journal.pone.0187644.
Gui, Y., Lei, X., and Huang, S. (2018) Collective effects of common SNPs and genetic risk prediction in type 1 diabetes. Clinical Genetics, 93: 1069-1074.
Lei, X., Yuan, D., and Huang, S. (2018) Collective effects of common SNPs and risk prediction in lung cancer. Heredity, doi:10.1038/s41437-018-0063-4.
Lei, X., Yuan, D., Zhang, Y., and Huang, S. (2018) Genetic continuity in the last seven millennia in human hepatitis B viruses. Communications in Information & Systems, 9: 357-373.
Zhang, Y. and Huang, S. (2019) Enrichment of de novo mutations in non SNP sites in autism spectrum disorders and an empirical test of the neutral DNA model. Communications in Information & Systems, 19: 343–355. DOI: https://dx.doi.org/10.4310/CIS.2019.v19.n3.a6.
He, P., Chen, N., et al. (2018) Neolithic super-grandfather Y haplotypes, their related surnames, and autism spectrum disorder. bioRxiv, doi: https://doi.org/10.1101/077222.
Yuan, D. and Huang, S. (2018) On The Peopling Of The Americas: Molecular Evidence For The Paleoamerican And The Solutrean Models. bioRxiv. doi: https://doi.org/10.1101/130989.
Zhang, Y., Lei, X., et al. (2018) Ancient DNAs and the Neolithic Chinese super-grandfather Y haplotypes. bioRxiv, doi: https://doi.org/10.1101/487918.
Zhang, Y. and Huang, S. (2019) The Out of East Asia model versus the African Eve model of modern human origins in light of ancient mtDNA findings. bioRxiv. doi:10.1101/546234.
Zhang, Y. and Huang, S. (2019) 古DNA的新发现支持现代人起源于东亚说. 人类学学报, 38: 491-498.
Liu, T. and Huang, S. (2019) Testing the basic tenet of the molecular clock and neutral theory by using ancient proteomes. bioRxiv, doi:10.1101/821736.
Chen, H., Lei, X., Yuan, D., and Huang, S. (2020) The relationship between the minor allele content and Alzheimer's disease. Genomics, doi:10.1016/j.ygeno.2020.01.015.
Chen, H., Zhang, Y., and Huang, S. (2020) Ancient Y chromosomes confirm origin of modern human paternal lineages in Asia rather than Africa. bioRxiv, doi:10.1101/2020.03.10.986042.
Chen, H. and Huang, S. (2020) Modern alleles in archaic human Y chromosomes support origin of modern human paternal lineages in Asia rather than Africa. arXiv, arXiv:2003.13473.
Wang, M., Wang, D., Yu, J., and Huang, S. (2020) Enrichment in conservative amino acid changes among fixed and standing missense variations in slow evolving proteins. PeerJ, 8:e9983. https://doi.org/10.7717/peerj.9983.
Zhu, Z., Lu, Q., et al. (2015) Collective effects of common SNPs in foraging decisions in Caenorhabditis elegans and an integrative method of identification of candidate genes. Sci. Rep., 5, 16904; doi: 10.1038/srep16904.
Zhu, Z., Lu, Q., et al. (2015) Compatibility between mitochondrial and nuclear genomes correlates with the quantitative trait of lifespan in Caenorhabditis elegans. Sci. Rep., 5, 17303; doi: 10.1038/srep17303.
Luo, D. and Huang, S. (2016) The genetic equidistance phenomenon at the proteomic level. Genomics, 108: 25-30. doi:10.1016/j.ygeno.2016.03.002.
Huang, S. (2016) New thoughts on an old riddle: what determines genetic diversity within and between species. Genomics, 108: 3-10. doi:10.1016/j.ygeno.2016.01.008.
Huang, S. (2016) Editorial: Special issue on the comprehensive functionality of genomic DNAs. Genomics, 108: 1-2. doi:10.1016/j.ygeno.2016.06.001.
Zhu, Z., Lu, X., et al. (2016) Close genetic relationships between a spousal pair with autism-affected children and high minor allele content in cases in autism-associated SNPs. Genomics, 109: 9-15.
Yuan, D., Lei, X., et al. (2017) Modern human origins: multiregional evolution of autosomes and East Asia origin of Y and mtDNA. bioRxiv. doi:10.1101/101410.
Yuan, D. and Huang, S. (2017) Genetic equidistance at the nucleotide level. Genomics, 109: 192-195.
He, P., Lei, X., et al. (2017) Accumulation of minor alleles and risk prediction in schizophrenia. Sci. Rep. doi:10.1038/s41598-017-12104-0.
Lei, X. and Huang, S. (2017) Enrichment of minor allele of SNPs and genetic prediction of type 2 diabetes risk in British population. PLoS ONE, 12(11): e0187644. https://doi.org/10.1371/journal.pone.0187644.
Gui, Y., Lei, X., and Huang, S. (2018) Collective effects of common SNPs and genetic risk prediction in type 1 diabetes. Clinical Genetics, 93: 1069-1074.
Lei, X., Yuan, D., and Huang, S. (2018) Collective effects of common SNPs and risk prediction in lung cancer. Heredity, doi:10.1038/s41437-018-0063-4.
Lei, X., Yuan, D., Zhang, Y., and Huang, S. (2018) Genetic continuity in the last seven millennia in human hepatitis B viruses. Communications in Information & Systems, 9: 357-373.
Zhang, Y. and Huang, S. (2019) Enrichment of de novo mutations in non SNP sites in autism spectrum disorders and an empirical test of the neutral DNA model. Communications in Information & Systems, 19: 343–355. DOI link.
He, P., Chen, N., et al. (2018) Neolithic super-grandfather Y haplotypes, their related surnames, and autism spectrum disorder. bioRxiv, doi:10.1101/077222.
Yuan, D. and Huang, S. (2018) On The Peopling Of The Americas: Molecular Evidence For The Paleoamerican And The Solutrean Models. bioRxiv, doi:10.1101/130989.
Zhang, Y., Lei, X., et al. (2018) Ancient DNAs and the Neolithic Chinese super-grandfather Y haplotypes. bioRxiv, doi:10.1101/487918.
Zhang, Y. and Huang, S. (2019) The Out of East Asia model versus the African Eve model of modern human origins in light of ancient mtDNA findings. bioRxiv, doi:10.1101/546234.
Zhang, Y. and Huang, S. (2019) 古DNA的新发现支持现代人起源于东亚说. 人类学学报, 38: 491-498.
Liu, T. and Huang, S. (2019) Testing the basic tenet of the molecular clock and neutral theory by using ancient proteomes. bioRxiv, doi:10.1101/821736.
Chen, H., Lei, X., Yuan, D., and Huang, S. (2020) The relationship between the minor allele content and Alzheimer's disease. Genomics, doi:10.1016/j.ygeno.2020.01.015.
Chen, H., Zhang, Y., and Huang, S. (2020) Ancient Y chromosomes confirm origin of modern human paternal lineages in Asia rather than Africa. bioRxiv, doi:10.1101/2020.03.10.986042.
Chen, H. and Huang, S. (2020) Modern alleles in archaic human Y chromosomes support origin of modern human paternal lineages in Asia rather than Africa. arXiv, arXiv:2003.13473.
Wang, M., Wang, D., Yu, J., and Huang, S. (2020) Enrichment in conservative amino acid changes among fixed and standing missense variations in slow evolving proteins. PeerJ, 8:e9983. https://doi.org/10.7717/peerj.9983.
Huang, S. (2019) Evolution, prime numbers, and an algorithm for the creative process. mathscidoc:1912.13001.
Xia, Z., Chen, H., Zhang, Y., and Huang, S. (2021) Ancient uniparental DNAs in distinguishing the competing theories of molecular evolution and modern human origins. Research Square, https://doi.org/10.21203/rs.3.rs-1087730/v1.
Wang, M. and Huang, S. (2023) The collective effects of genetic variants and complex traits. J. Human Genetics, 68: 255–262.
Huang, S. (2023) The maximum genetic diversity theory of molecular evolution. Communications in Information and Systems, 23: 359-392. https://dx.doi.org/10.4310/CIS.2023.v23.n4.a1.
Huang, S. (2023) Examining models of modern human origins through the analysis of 43 fully sequenced human Y chromosomes. bioRxiv, https://doi.org/10.1101/2023.11.09.566475.
Zhu, Z., Han, C., and Huang, S. (2024) New insights shed light on the enigma of genetic diversity and species complexity. Sci China Life Sci, 67: 2774–2776.
Huang, S. (2025) Reconstructing the Huang Surname and Its Related Lineages: A Comprehensive Analysis of Molecular Genetics and Historical Genealogies. bioRxiv, https://doi.org/10.1101/2025.02.17.638729.
Huang, S. (2025) A Rebuttal to Zhang’s Critique of the Genetic Equidistance Phenomenon and Maximum Genetic Diversity Hypothesis. ChinaXiv, https://doi.org/10.12074/202502.00109.
Huang, S. (2025) A Rebuttal to Zhang's Critique of the Genetic Equidistance Phenomenon and Maximum Genetic Diversity Hypothesis. Zenodo, https://doi.org/10.5281/zenodo.14927073.
Huang, S. (2025) The Maximum Genetic Diversity Theory: A Comprehensive Framework for Understanding Evolutionary Processes. BioCosmos, 5: 41-61. https://doi.org/10.2478/biocosmos-2025-0009.
Huang, S. (2025) Revisiting Human Evolution: Insights from an Expanded Framework of Evolutionary Processes. Zenodo, https://zenodo.org/records/15333528.
Books
Discover comprehensive works by Shi Huang and others on the Maximum Genetic Diversity (MGD) theory and related topics:
Ho, M. W. (2010) Development and evolution revisited.
In Hood, K. E., Halpern, C. T., Greenberg, G., et al. (Eds.),
Handbook of Developmental Science, Behavior, and Genetics.
Malden, MA: Wiley Blackwell.
Denton, M. (2016) Evolution: still a theory in crisis.
Seattle, WA: Discovery Institute Press.
Huang, S. (2008) Histone methylation and the initiation of cancer.
In Tollefsbol, T. (Ed.), Cancer Epigenetics. CRC Books.
Huang, S. (2014) Human origins as told by informative DNAs.
In Xi, H., Liu, W., & Chen, Z. (Eds.),
The development of Chinese Anthropology in the 21st century.
Intellectual Property Press, Beijing.
(Chinese title: 用与种系发生相关的DNA序列解读人类起源)
Huang, S. and Zhu, Z. (2022) Systems Biology, a textbook.
East South University Press, Nanjing, China.
Martin, M. K. (2023) The wrong ape for early human origins:
the chimpanzee as a skewed ancestral model.
Lanham, MD: Lexington Books.
Bickel, D. (2022) Testing hypotheses of molecular evolution.
In Phylogenetic trees and molecular evolution.
Springer Briefs in Systems Biology. Cham: Springer; p. 71–77.
Note: This section will be updated as more books are published or identified.
Media Coverage
Explore how the Maximum Genetic Diversity (MGD) theory and the
"Out of East Asia" hypothesis are covered in the media:
Daily Mail
– Scientist challenges 'Out of Africa' theory with new origin for modern humans.
South China Morning Post
– It was East Asia: Chinese scientist challenges ‘Out of Africa’ theory of human evolution.
Learn about the background and expertise of Dr. Shi Huang, the originator of the
Maximum Genetic Diversity (MGD) theory.
Education
9/84 – 12/88: Ph.D., Biochemistry, University of California, Davis, CA
9/79 – 7/83: B.S., Genetics, Fudan University, Shanghai, China
10/61 – 9/79: Grew up in the army compound of the Academy of Military Medicine
in Beijing, where his father served as a professor. Enjoyed sports, mathematics,
and the arts. Received formal extracurricular art training at the Teens’ Palace
and took part in the 1978 entrance examination for the Central Academy of Art.
Professional Experience
2009 – Present: Professor, Furong Scholar, Department of Genetics, Central South University
7/98 – 12/08: Associate Professor, The Sanford-Burnham-Prebys Medical Discovery Institute, La Jolla, California
4/92 – 6/98: Assistant Professor, The Sanford-Burnham-Prebys Medical Discovery Institute, La Jolla, California
1/89 – 4/92: Postdoctoral Fellow, University of California, San Diego
9/84 – 12/88: Graduate Student, University of California, Davis, CA
Awards and Honors
1993 – 1997: Pew Scholar in the Biomedical Sciences
1991 – 1992: American Heart Association Postdoctoral Fellowship
1990 – 1991: Fight for Sight Fellowship
1989 – 1990: CRCC Postdoctoral Fellowship
1985 – 1988: Distinguished Scholar Fellowship, UC Davis
1984 – 1985: CUSBEA Fellowship, Department of Education, China
Frequently asked questions about the Maximum Genetic Diversity (MGD) theory and related topics:
What is the MGD theory?
The MGD theory, developed by Dr. Shi Huang, posits that genetic diversity reaches an upper
limit determined by species complexity, challenging the neutral theory and supporting the
"out of East Asia" model of human origins. It reinterprets the
GEP (Genetic Equidistance Phenomenon) to suggest natural selection maintains this maximum.
Does maximum genetic diversity mean a stop of evolution?
No. Mutation followed by either drift or selection will always occur.
Can the neutral theory be called a no-MGD theory?
Yes.
How does the 'out of East Asia' hypothesis differ from 'out of Africa'?
The 'out of East Asia' hypothesis, based on MGD, argues that modern humans originated in
East Asia, supported by ancient DNA showing closer genetic ties to Asians than Africans.
The 'out of Africa' model claims a southern African origin around 50,000 years ago, based on
the debunked molecular clock hypothesis and the neutral theory. It misinterprets the higher
genetic diversity of sub-Sahara Africans as still scaling with time rather than already being
at the upper limit level.
How do you refute the bottleneck hypothesis of the out of Africa theory?
The bottleneck hypothesis proposes that the lower genetic diversity of non-Africans,
compared to Africans, can be explained by a near-extinction event affecting the groups
that left Africa for Eurasia. This idea can be challenged on several grounds:
Contrary real-world examples: Populations known to have undergone severe
bottlenecks, such as the indigenous peoples of Australia and the Americas, actually
exhibit higher genetic diversity than East Asians. East Asians, despite being the
largest population on Earth and showing no evidence of past bottlenecks, have the lowest
genetic diversity.
Pattern of fixed alleles: East Asians have more fixed derived alleles,
particularly in fast-evolving genes. Bottlenecks should not preferentially fix
fast-evolving alleles over slow-evolving ones.
Variant-type diversity differences: The diversity gap between Africans
and non-Africans is much larger for fast-evolving variants than for slow-evolving
variants, which is inconsistent with the expected uniform diversity reduction from a
bottleneck.
Is the MGD theory widely accepted?
The MGD theory is innovative and controversial, facing resistance from mainstream academia
due to its challenge to the neutral theory. However, it is gaining attention, as seen in
media coverage and ongoing research, though it requires further validation.
Recent Updates — Top 10 Monthly Developments for September 2025
The Yunxian Man advances the independent branch of Homo sapiens to at least 1 million years ago, supporting the Out of East Asia hypothesis's conclusion of 2 million years ago and falsifying the Out of Africa hypothesis's conclusion of 600,000 years ago.
Read more
Denisovans are Africans! A PNAS paper found that Denisovans were phenotypically archaic Africans (Kabwe), confirming our conclusion using informative DNA in our 2017 preprint.
Read more
Huangshi submitted an invited paper titled "Research on the Origin of Homo sapiens and Civilization History in the Transformation of Molecular Evolution Theory" to the first "New Debate on Ancient History and the Revival of Chinese Civilization" conference, and it received coverage from official media.
WeChat,
VOC News
Overall increase in somatic mutations in short housekeeping genes is associated with decreased gene expression during brain aging. This is unexpected if most mutations are neutral.
Read more
mtDNA variants are not neutral. Reversible compromise of physiological resilience by accumulation of heteroplasmic mtDNA mutations.
Read more
Genome size varies enormously, and while neutralists have long assumed this variation to be functionally irrelevant, growing evidence now points to important adaptive roles. Genome size variation is attributed to adaptive purging of transposable elements.
Read more
A rare paper using ancient DNA to verify neutral theory predictions of genetic distance, with ambiguous results, mostly falsifying it, but the method has flaws. Ancient host-associated microbes obtained from mammoth remains.
Read more
Coding regions of a gene encode not just protein sequences but also regulatory functions controlling its own gene expression. Not a finding friendly to the neutral theory.
Read more
A recent review published domestically promoting the Out of Africa hypothesis continues to repeat the usual tactics of Western leftist patriarchs, selectively discussing issues, firmly avoiding mention that the neutral theory is the theoretical premise of the Out of Africa hypothesis. The only time it mentions it is a brief introduction to the Out of East Asia hypothesis, noting Huangshi's critique of the neutral theory and the Out of Africa hypothesis. The believers of the Out of Africa hypothesis shyly dare not mention the neutral theory, which can only have one explanation: it's too rotten and really can't hold up on stage.
Read more